Our Products Cannot Be Used As Medicines Directly For Personal Use.


Welcome! For price inquiries, please feel free to contact us through the form on the left side. We will get back to you as soon as possible.
TdT
| Catalog | Product Name | EC No. | CAS No. | Source | Price |
|---|---|---|---|---|---|
| NATE-1926 | Terminal Transferase from Calf, Recombinant | EC 2.7.7.31 | 9027-67-2 | E. coli | Inquiry |
| EXWM-3242 | DNA nucleotidylexotransferase | EC 2.7.7.31 | 9027-67-2 | Inquiry | |
| NATE-0692 | Native Calf Terminal Transferase | EC 2.7.7.31 | 9027-67-2 | Calf thymus | Inquiry |
Related Reading
TdT (EC 2.7.7.31, Terminal deoxynucleotidyl Transferase) belongs to the pol X family of DNA polymerase. TdT is the first DNA polymerase identified in mammals and purified from calf thymus extract in 1960. The accepted name of this enzyme is DNA nucleotidylexotransferase. Other names in common use include terminal addition enzyme, terminal deoxynucleotidyltransferase, terminal deoxyribonucleotidyltransferase, and terminal transferase. TdT uses only single-stranded DNA as a nucleic acid substrate, incorporating nucleotides in a template-independent manner to complete DNA synthesis. Upon V(D)J recombination, TdT randomly adds nucleic acids to single-stranded DNA. Through the delicate randomization of genetic material, TdT plays an important role in the evolution and adaptation of the vertebrate immune system. The ability of TdT to randomly incorporate nucleotides contributes to the production of different immunoglobulin and T cell antigen receptors, increasing the diversity of antigen receptors.
Structure
The structure of TdT is similar to that of all X family DNA polymerases, with a Polβ-like domain at the C-terminus, including thumb, fingers, plam and 8 kDa domains. TdT also has a "lariat-like" loop that prevents interaction between the enzyme and the template strand. In addition, the 8kDa domain, also known as the index finger domain, contacts the thumb domain to form a cavity that allows dNTPs to diffuse to the active site of the enzyme. This feature defines the general form of TdT as a ring structure in which the primer is perpendicular to the axis of the cavity on the palm domain. In addition to the Polβ-like domain, the N-terminus of TdT also includes a nuclear localization signal (NLS) motif and the breast cancer susceptibility protein BRAC1 C-terminal (BRCT). The BRCT domain mediates protein/protein and protein/DNA interactions in the DNA repair pathway and cell cycle checkpoint regulation following DNA damage. The BRCT domain in TdT interacts with Ku70/86, a protein heterodimer that recognizes and binds to free DNA ends during V(D)J recombination and double-strand break repair.
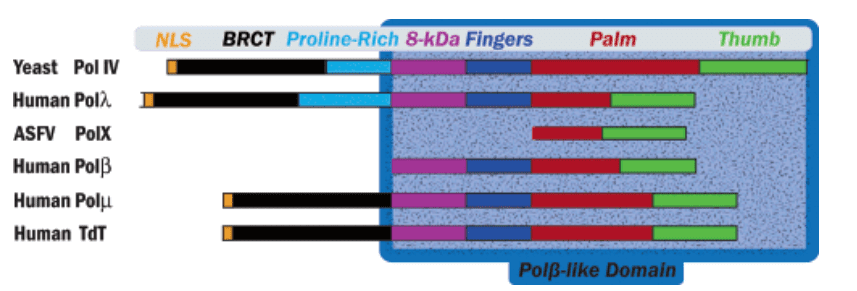 Figure 1. Domain organization of X-family DNA polymerases. (Fowler J.D. 2006)
Figure 1. Domain organization of X-family DNA polymerases. (Fowler J.D. 2006)
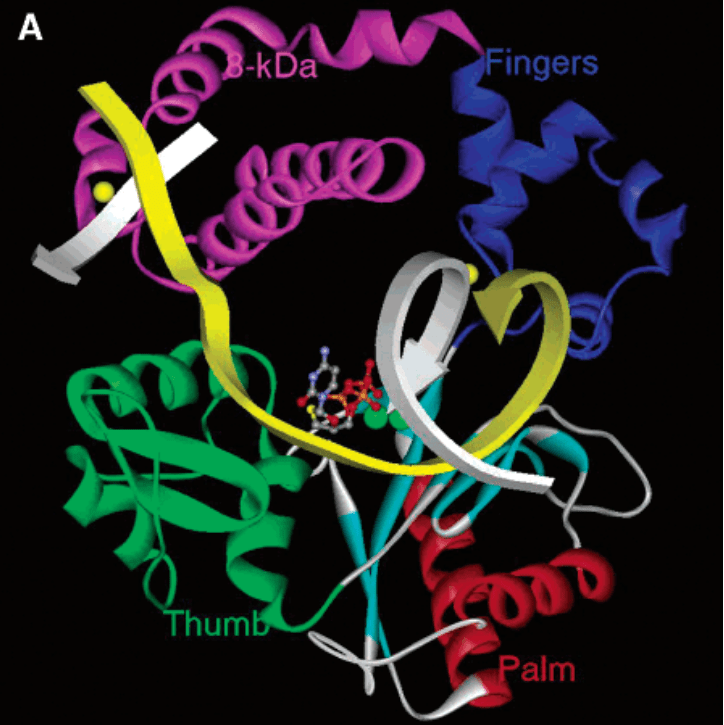 Figure 2. Structure of Polβ-like domain. (Fowler J.D. 2006)
Figure 2. Structure of Polβ-like domain. (Fowler J.D. 2006)
Catalytic Mechanism
A striking feature of the X-family DNA polymerase is that the catalytic site is highly conserved, with three strictly conserved Aspartates coordinating two essential metal ions that participate in the two-metal ions mechanism. Metal A activates the 3'OH of the last nucleotide to allow the attack of α phosphate while metal B enters to stable leaving group (PPi) along with the nucleoside triphosphate. In TdT, the coordination geometry of divalent metal ions, including transition metal ions such as Mn++, Co++ and Zn++, is studied in atomic detail during the fully catalytic site, and their activity on nucleotidyltransferase is more efficient than Mg++. In order to translocate the newly extended primer strand to the catalytic site for a new addition, metal A must exit and be replaced by Na+.
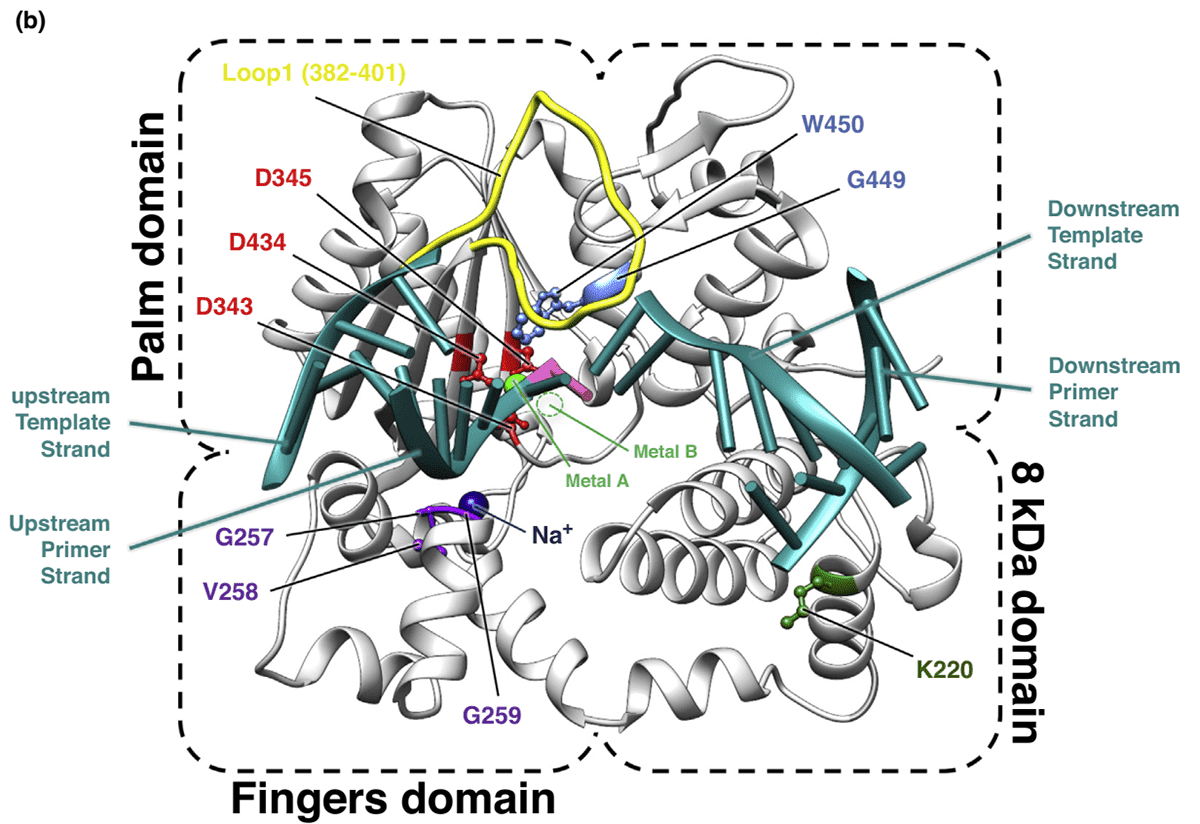 Figure 3. Mechanism of TdT bound to a DNA synapsis. (Loc’h J. 2018)
Figure 3. Mechanism of TdT bound to a DNA synapsis. (Loc’h J. 2018)
TdT can add random deoxyribonucleotides (dNTPs) to ssDNA primers in a template-independent manner, with primers of at least 3 nucleotides in length. In vitro experiments have shown that TdT can use four natural dNTPs, but TdT preferentially incorporates dCTP and dGTP compared to dATP and dTTP. Moreover, the TdT nucleotide binding site can accommodate deoxyribonucleotides and ribonucleoside triphosphates (dNTPs and rNTPs).
Application
TdT can be used as an anti-cancer target. Clinical evidence suggests that changes in TdT activity and expression levels play an important role in cancer initiation, progression, and chemotherapy response. In B-cell and T-cell acute lymphoblastic leukemia (ALL) and acute myeloid leukemia (AML), TdT expression is elevated, and high levels of TdT activity are associated with poor prognosis. Studies have shown that TdT-positive patients have a two-fold reduction in remission rate compared with TdT-negative leukemia patients. These findings have driven research into selective inhibitors of TdT that act as chemotherapeutic agents against these forms of leukemia.
TdT can be used as a biochemical tool. TdT can utilize a variety of nucleotide analogs, and TdT is tolerant to a large number of modifications of nucleases, which provides an efficient method in labeling the break of double-stranded DNA in vivo and in vitro. TUNT (TdT-mediated dUTP-biotin nick end-labeling) is a TdT-based technology that efficiently incorporate biotinylated dUMP to the 3' end of single-stranded DNA at the DNA break site. The bound biotin dUMP can be detected by fluorescent labeling and can directly quantify the number and location of DNA breaks. This technique is commonly used to detect apoptosis and programmed cell necrosis in eukaryotic cells. TdT can also be used as a biocatalyst to label the 3’ end of a synthetic oligonucleotide with a radionucleotide or fluorescent probe. These labeled primers are annealed to a complementary strand and can be used as a radioactive substrate to detect the activity of an enzyme in nucleic acid metabolism.
References
- Fowler, J.D., Suo, Z. Biochemical, structural, and physiological characterization of terminal deoxynucleotidyl transferase. Chem. Rev, 2006, 106:2092−2110.
- Loc’h, J., Delarue, M. Terminal deoxynucleotidyltransferase: the story of an untemplated DNA polymerase capable of DNA bridging and templated synthesis across strands. Curr Opin Struct Biol, 2018, 53:22-31.
- Motea, E.A., Berdis, A.J. Terminal deoxynucleotidyl transferase: the story of a misguided DNA polymerase. Biochim Biophys Acta, 2010, 1804(5): 1151–1166.